酵母は invertase を細胞外に分泌して蔗糖を分解し、分解された糖は誰でも利用できる—ここが面白いところだ。細胞は「裏切り」を選べる:隣が分泌した酵素にただ乗りして、自分では分泌しない。研究者は機能的な SUC2 遺伝子を持つ酵母を「協力者」、SUC2 を欠失させた酵母を「裏切り者」と呼び、両者を競争させた。
結果は直感に反する。
- 疎な集団(社会的密度が低い)では、裏切り者の fitness はわずか 0.87—協力者より劣る。
- 密な集団(社会的密度が高い)では、裏切り者の fitness は 1.19 まで上がる—協力者を上回る。
なぜか。社会的密度が高くなるほど、協力者は他の協力者と出会いやすくなる。皆で酵素を分泌すれば、公共財の規模が大きくなり、全員の利得が上がる。そこへ裏切り者が紛れ込み、コストを支払わずに公共財を一方的にもうけ、利得が跳ね上がる。
密度がきわめて高くなると、裏切り者はほぼ常に標的を見つけ、自分で分泌するより多くを盗める。協力者は逆に足を引っ張られる。
これは私の直感にしっくりくる。ただ、自分で一度走らせてみたかった—論文の結論を検証するためではなく、この過程を自分の目で「見る」ため、方程式を格子に書き込み、数字が走るのを見届けるために。
モデル設定
$n \times n$ の格子に、population density が各セルに agent を配置する確率を制御する。Agent には 2 種類の戦略がある:C(協力、酵素を分泌する)と D(裏切り、分泌しない)。
二者が出会うと、標準的な囚人のジレンマをプレイする。payoff 行列は以下のとおり:
$$ \begin{pmatrix} R=3 & S=0 \\ T=5 & P=1 \end{pmatrix} $$R は相互協力の報酬、T は裏切りの誘惑、S は搾取される側の利得、P は相互裏切りの罰。古典的な設定に従い:$T > R > P > S$、かつ $2R > T + S$(相互協力は繰り返しの裏切りより優れている)。
各ステップ:
- Agent は Von Neumann 近傍(上下左右の 4 セル)で隣を探す。
- 二者が 1 局プレイする。
- このステップの得点 $\pi$ を比較する。
- 利得差に比例する確率で隣人の戦略を模倣する。
- ランダムな方向に 1 セル移動する。
見るのは当期の得点だけ。履歴は残さず、計画も立てない。
理論的予測
複製者動学(Replicator Dynamics)は平均場の方程式を与える:
$$\frac{dx}{dt} = x(1-x)[\pi_C - \pi_D]$$$x$ は協力者の割合、$\pi_C$ と $\pi_D$ は二つの戦略の期待単ステップ利得である。
利得は出会い確率に依存する。平均場近似では、協力者が協力者と出会う確率は $x$、裏切り者と出会う確率は $1-x$。裏切り者はその逆となる:
$$\pi_C = x \cdot R + (1-x) \cdot S$$$$\pi_D = x \cdot T + (1-x) \cdot P$$代入すると:
$$\pi_C - \pi_D = x(R - T) + (1-x)(S - P)$$具体値 $R=3, S=0, T=5, P=1$ を用いると:
$$\pi_C - \pi_D = x(3-5) + (1-x)(0-1) = -2x -1 + x = -1 - x$$$\pi_C - \pi_D < 0$ は常に成立する—$x$ の大きさに関わらず、協力者の当期利得は常に裏切り者より低い。
したがって $\frac{dx}{dt} = x(1-x)(-1-x) < 0$ となる。協力者の割合は単調に減少し、最終的に $x=0$(完全裏切り)へ向かう。
平均場近似は「十分混合」された世界を仮定する:すべての個体はすべての他個体と等確率で出会う。しかし私の格子はそうではない。空間構造が協力を支えてくれるかもしれない。
シミュレーションと結果
以下の実験は $50 \times 50$ の格子上で実行し、各ステップで同期更新し、系が落ち着くまで十分に多くのラウンドをプレイする。
実験 1:密度を固定し、進化軌道を観察する
population density $\rho = 0.3$ に設定し、初期協力者割合 $x_0$ を変化させる。
直感的な予想:$x_0$ が高ければ高いほど、協力者は長く持ちこたえる。低い $x_0$ はすぐに崩壊する。だが格子構造により、局所的に協力率の高い領域が初期段階でしばらく持ちこたえる可能性がある。
実際に走らせてみると、協力者の「生存窓」はきわめて狭い。空間構造が局所的クラスタリングによって短い防御期間を引き延ばすものの、D 戦略はやがて境界から浸透してくる—協力クラスターの周縁部は常に最初に侵食される。
実験 2:初期割合を固定し、密度効果を観察する
$x_0 = 0.5$ に設定し、population density $\rho \in [0.02, 1.0]$ を変化させ、ラン終了後に $\pi_C$ と $\pi_D$ の平均値の差を算出する。
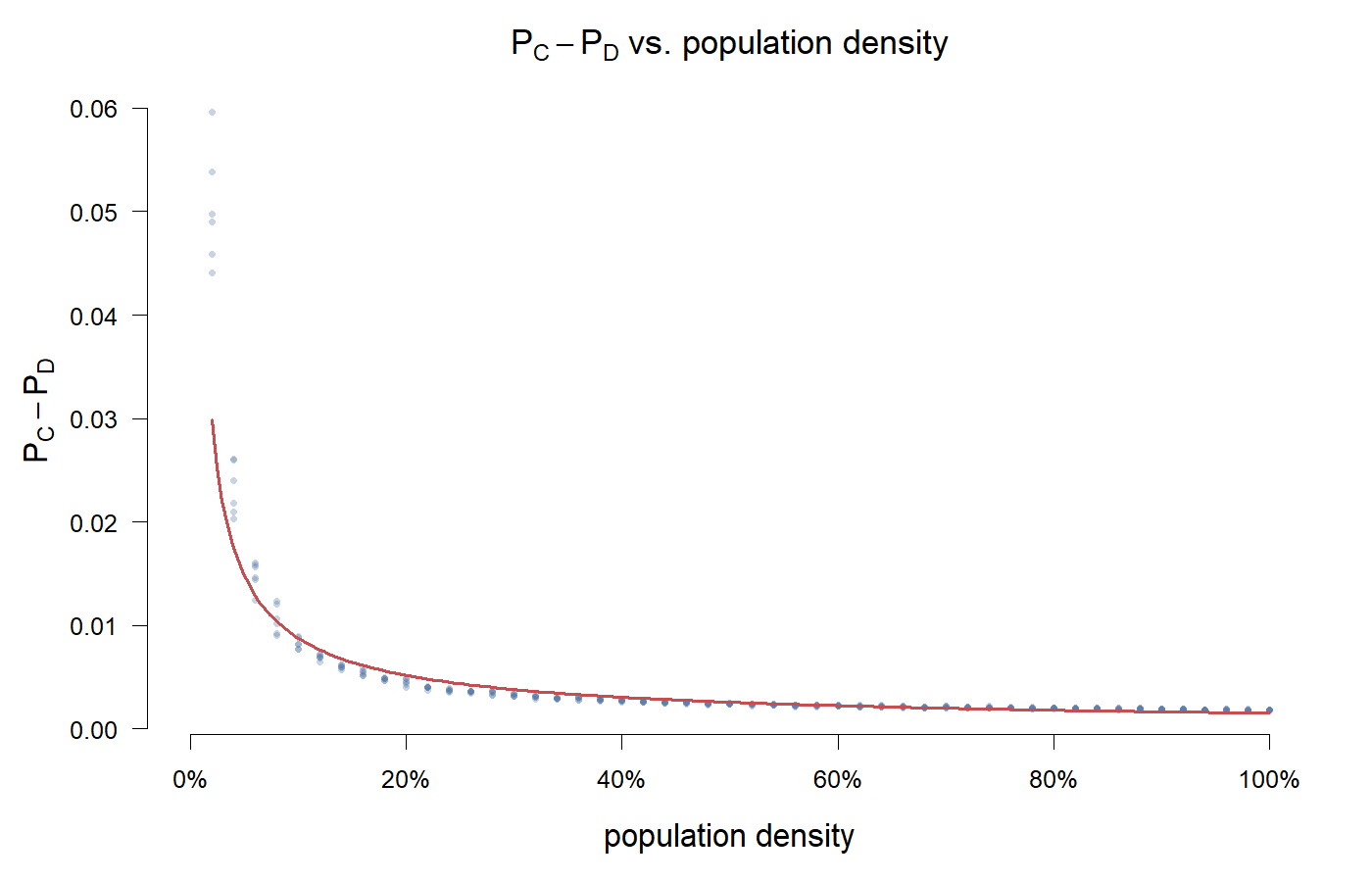
縦軸は $P_C - P_D$、すなわち協力者の利得から裏切り者の利得を引いた値である。
しかし今回は、グラフに大きな欠落がある—高密度領域はすべて空白になっている。プロットミスのためではなく、測定不能だからだ。
理由は単純:$P_C - P_D$ は両戦略が生き残っていてこそ意味を持つ。burn-in 100 ステップ経過後から計測を始めたが、高密度下では C は 100 ステップも生き残らない—6 ステップ目で絶滅する。$\pi_C$ は NaN であり、欠損値ではなく「測定できる協力者がいない」という意味だ。適当な数字を埋めるのは自ら欺くようなものだ。
測定可能な範囲($\rho < 0.05$)では、$P_C - P_D$ は一貫して負であり、-0.03 から -0.16 まで下落する。協力者の平均利得は裏切り者を下回る。これは複製者動学の予測と一致する:$\pi_C - \pi_D = -1 - x < 0$。空間構造は、測定可能な範囲では符号を反転させなかった。
密度 2% では格子がまばらで、agent の大部分は孤立している。孤立した agent はペアになる相手がいない—ゲームなし、利得の変動なし、現戦略の維持は損にならない。協力者が生き残るのは「互恵の優位性」のためではなく、孤立によって戦略が凍結されるためだ—D はそもそも彼らに接触できない。時折遭遇すれば、D の当期利得は C を上回り、模倣確率は D に傾き、C はじわじわと失われていく。
密度が高くなるほど、遭遇は頻発する。出会うたびに C は D に搾取される機会を得る。C-C の互恵がより多く起きても、両者がコストを払って公共財を蓄積する—まさに何の貢献もしない隣の D を補助する形になる。
Greig の酵母実験も同じ方向を示している:密度が高ければ高いほど、裏切りは有利になる。ただ彼らの結果は裏切り者の観点から見たものだ—高密度下では裏切り者の fitness > 1。私のプロットは協力者の観点から見たものである—測定可能な範囲では、不利の度合いは密度とともに増す。高密度領域で何が起きるかは、このプロットから読み取ることはできない。
興味深いことに、このパラメータ設定では臨界密度は一度も現れなかった—ゼロに到達しなかったからではなく、ゼロに到達する余地がなかったからだ。C は臨界点に届く前に死に絶えた。より極端な payoff 設定(例えば T をより高くする)であれば、このわずかな測定可能範囲すら消えるかもしれない。
(あるいは Von Neumann を Moore 近傍に置き換える—4 近傍より 8 近傍のほうが、D に曝されやすくなる。まだ試せていない。)
実験 3:協力者の存続 vs 密度
実験 2 では答えられなかった問いに、実験 3 が応える。定常状態にも burn-in にも頼らない—C がどれほど持ちこたえるかを直接追跡する。
$x_0 = 0.5$ に固定し、$\rho \in [0.02, 1.0]$ を変化させ、200 ステップ走らせて、3 つの量を記録する:時間平均の協力者割合 $\bar{x}$、最終ステップの協力者割合 $x_{\text{final}}$、初回絶滅ステップ $t_{\text{collapse}}$。
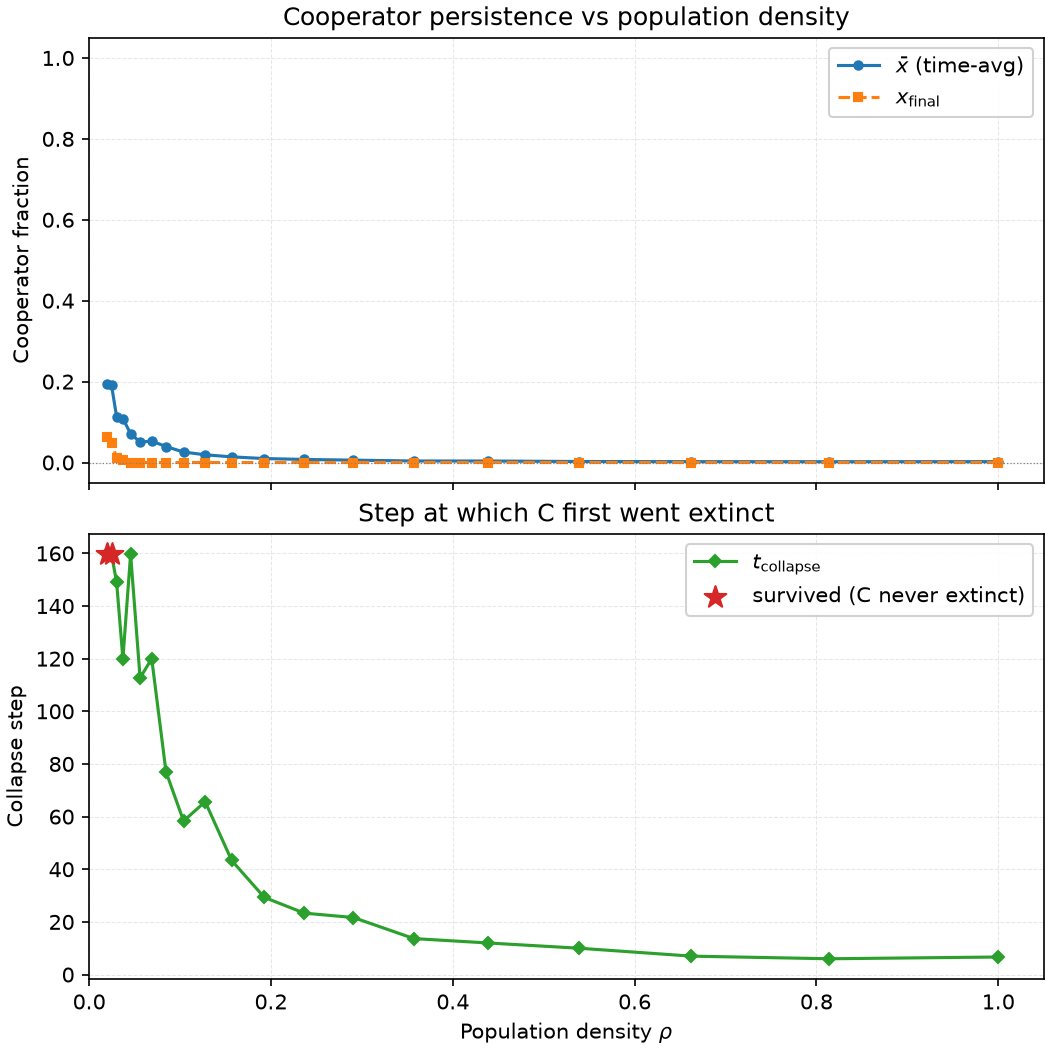
3 つの密度区間、3 つの運命:
- 低密度($\rho < 0.03$):$t_{\text{collapse}} = \text{NaN}$—C は 200 ステップ以内に一度も絶滅しない。しかし $\bar{x}$ は 0.11〜0.19 にとどまり、$x_{\text{final}}$ は 0.01 まで下がる。C が生き残るのは強さ故ではなく、孤立故である—agent の大部分は隣がいない時間が長く、ゲームも模倣も起きず、戦略が凍結される。接触できない C を D は侵食できない。
- 中密度($\rho \in [0.03, 0.44]$):$t_{\text{collapse}}$ は 160 ステップから 12 ステップへ単調に減少する。密度が高くなるほど、D は搾取対象の C を見つけやすくなり、C のクラスターはより速く浸透・解体される。$\bar{x}$ は 0.11 から 0.004 へ落ちる—協力の総量は密度とともに減少する。
- 高密度($\rho > 0.54$):$t_{\text{collapse}} \approx 6$〜$10$、$\bar{x} \approx 0.002$。十分混合近似が成立し、D は厳密に支配的となり、C は 10 ステップ以内に除去される。
実験 2 は $\rho \geq 0.06$ で完全に測定不能になる—C は burn-in 前に死に絶える。実験 3 がこの空白を埋める。
その後の考察
Greig の論文から、一つのことに気づかされた:囚人のジレンマの「ジレンマ」は、解けないことにあるのではない—その解が構造に依存していることにある。同じゲームのルールが、疎なソーシャルネットワークと密なソーシャルネットワークで逆方向の結果を生む。
複製者動学は基準となる予測を提供する—十分混合・当期利得のみを見た場合の進化方向だ。しかし現実の系の空間構造—寒天培地上の近傍半径であれ、格子上の Von Neumann 近傍であれ—それを組み込むと、予測は変わる。
ABM の価値はここにある:構造を入れ、何が創発するかを見る。
実験 1〜3 を走らせた感覚では、空間構造は緩衝材のように働く—方向を反転させるのではなく、速度を遅らせるのだ。協力者は D に負ける運命にあるが、格子の上では平均場方程式の予測より遅く負ける。低密度では「崩壊しない」まで遅延する(凍結されているだけだが)、中密度では数十〜数百ステップ遅延し、高密度では 6 ステップしか遅延しない。
だが書き終えてから、考えれば考えるほど、物足りなさを感じた。
Greig の実験は蔗糖プレート上で行われた。蔗糖は唯一の炭素源である。疎なプレートでは糖が多く細胞が少ない—各細胞周囲の蔗糖はほぼ無限とみなせる。密なプレートでは細胞が密集し、糖は急速に消費される—自分の周囲の糖は、隣がどれだけの酵素を分泌したかに依存する。
私の ABM は、payoff 行列が固定だと仮定している。$T=5, R=3, P=1, S=0$—密度に関わらず、一度のゲームの利得は永遠に変わらない。しかし酵母の invertase は、環境と切り離された固定的な payoff 関数ではない。蔗糖を分解する。糖が減れば分解量も減る。分解量は、自分が酵素を分泌するかどうかに加えて、蔗糖がどれだけ残っているかにも依存する。
実験 3 はこの差を鮮明に示す:高密度下では C は 6 ステップで絶滅する。しかし Greig の実験では、高密度下でも協力者は存続している(fitness 0.87〜1.19)。固定行列ではこれを説明できない。もし C がこれほど速く消えるなら、Greig の酵母集団はとうの昔にすべて裏切り者になっているはずだ—しかし現実はそうなっていない。
蔗糖が尽きれば—密集した細胞群が誰も酵素を分泌しないなら—相互裏切りの payoff は $P=1$ ではなく、負の値になるかもしれない。何も分解されない。細胞は飢える。逆に C-C は酵素コストを支払う必要があるが、少なくともいくらかの糖は得られる。
つまり、Greig の酵母は常に囚人のジレンマをプレイしているとは限らないのかもしれない。
糖が豊富な高密度領域では、裏切り者は協力者を搾取して満腹になる—典型的な PD。しかし糖が枯渇した縁辺部—疎な群落が局所の蔗糖を使い果たした後—ゲームの構造は囚人のジレンマから別のものへと移行する。相互裏切りはもはや低利得ではなく、負利得となる。協力の相対的な不利は突然消える。
戦略は資源を消費し、資源は行列を書き換え、行列は戦略を再形成する。このループが一度閉じると、囚人のジレンマの「ジレンマ」は、与えられた境界条件を持つ最適化問題ではなくなる—その境界条件自体が変数なのだ。
実験 3 の $t_{\text{collapse}} \approx 6$ は終点ではなく、起点である—固定行列下では協力者があまりにも速く死に、生物学的実験と食い違うことを示している。この仮説を検証するには、酵素動学を ABM に組み込む必要がある—糖濃度を局所変数とし、payoff を糖濃度からリアルタイムで算出する。「密度が payoff 行列を決定する」のではなく、「密度が資源消費を通じて payoff 行列を間接的に変化させる」。
一つの方向。
参考文献
Greig, D., & Travisano, M. (2004). The Prisoner's Dilemma and polymorphism in yeast SUC genes. Proceedings of the Royal Society of London. Series B: Biological Sciences, 271, S25 - S26.