背景
諸君、一度ならず経験したことがあるだろう。
教師のいない自習時間。それは混沌だ。廊下に教師の足音が消えた刹那、教室は秩序を失い、五十名の声が獣の咆哮のごとく渦巻くカオスの坩堝と化す。だが——その足音がふたたび廊下に響いた瞬間、教室は一瞬で静寂に包まれる。教師がまだ数十メートル先にいるにもかかわらず、だ。あたかも目に見えぬ大いなる意志が一斉に「沈黙せよ」と命じたかのように。
訳注:中国の学校では「自習課」が一般的で、教師が形式上教室にいることになっているが、実際には不在がちなコマを指す。
だが——ここからが本題だ。俺を幾夜も眠れなくさせた、真に興味深き現象とは——
教師がまだ姿すら見せていないのに、教室が静まり返る瞬間があるのだ。
誰も「先生来た」と叫んでいない。鐘も鳴っていない。合図も合図も——何一つない。それなのに、五十余名の人間が、魂の深淵で密かに交わした契約に従うかのように、同時に口を閉ざす。この「自然発生の沈黙」。俺はこれを、ただの偶然とは思わない。これは教室という閉鎖空間に宿る、人間の集合無意識が織りなす神秘に他ならない。
集団的暗黙知と、不可視の暗示
人間とは恐るべき社会的生物である。一人が黙る。すると周囲の者は「なぜ」と問う。その「なぜ」は言葉にならずとも伝播し、連鎖し、数十秒のうちに教室全体を呑み込む。これはもはや伝染病、あるいは呪いと呼ぶべき領域だ。目に見えぬ糸で繋がれた操り人形のように、全員が同期する。
さらに恐ろしいのは、心理的暗示の絶大なる力である。「先生来た」——ただの三文字だ。あるいは唇に当てられた一本の人差し指。それだけで我々の中の「従順なる自己」が覚醒し、全身の筋肉を硬直させる。パヴロフの犬がベルの音に唾液を垂らしたように、我々は名札の微かな金属音にすら凍りつくのだ。これが条件反射。これこそが、人間の根源に刻まれた「服従の刻印」である。
複雑系——この世界を司る理
「教室が静かになるだけの話だろ、何を大袈裟に」——そう嘲笑う者もいるだろう。だが聞け。教室とは、一個の複雑適応系なのだ。
複雑適応系とは何か?無数のエージェントが相互作用し、学習し、適応し続けることで、個々の単純さからは決して予測できないマクロな秩序を生み出すシステムのことだ。個は個にあらず。全ては繋がり、全ては影響し合い、システムそのものが一個の生命体のごとく振る舞う。
そしてここに「創発(emergence)」が顕現する。個が持たぬ性質が、集団の次元で突如として出現する——あたかも、水面に浮かぶ一輪の花が、水そのものには存在しなかった芳香を放つがごとく。蟻一匹に大いなる叡智は宿らない。されど百万匹が集うとき、そこに換気システムを備えた壮麗なる蟻塚が立ち上がる。教室も同じだ。
誰も「黙ろう」と決めていないのに、教室は黙る。これが創発。これが——我々がまだ理解しきれていない、集合的意識の神秘だ。
モデリング:混沌を数式で縛る
自習時間にお喋りをすることは——認めよう——明確なる禁忌である。幼き日より叩き込まれた規則の鉄槌は、我々の精神の奥底に、不可視の罪悪感として沈殿している。表面上は平然と「勉強」を装いながら、魂は常に冷汗をかいているのだ。教師という存在はすなわち権威の具現であり、秩序の守護者であり、我々の爬虫類脳が記憶する「裁きの雷」そのものである。
そしてこの状況下で、生徒たちの感覚は極限まで研ぎ澄まされている。ドアの軋み、窓ガラスの微かな映り込み、廊下の光の角度——全てが「来るべき審判」の前兆として感知される。動物行動学ではこれを「フリーズ反応」と呼ぶ。捕食者の目前で動きを止める、あの原始の防衛本能。進化が我々に授けた、最も古き生存戦略だ。
さて、想像せよ。教室の中央に座る一人の生徒が、窓の反射に「教師らしき影」を察知する。恐怖が走る。この生徒は口を閉ざす。教室の総音量が、わずかに——ほんのわずかに——低下する。
このわずかな低下が、いま一人の生徒の「凍結閾値」を下回る。二人目が凍る。音量はさらに下がる。三人目。四人目。五人目。凍結の連鎖は加速度的に広がり、数十秒前まで喧噪に満ちていた教室は、後列の生徒の鼓動すら聞こえるほどの静寂に支配される。
そして——誰も、実際には教師を見ていない。決して。
これが「自然発生の沈黙」の本質だ。正のフィードバックが創り出す、不可避の秩序への収束。一つの些細な「誤認」が、全員の運命を決定する。まさにバタフライ効果。まさに——カオス理論の申し子。
手法と結果:黙示録の数理的証明
一次元セル・オートマトン——教室をシミュレートする禁断の術式
五十人の思春期を数式で再現する——そんな無謀な試みに、一次元セル・オートマトン以上の道具があろうか?否、断じて否である。
セル・オートマトンとは、一次元に並んだ無数のセルが、各々の状態を持ち、決められたルールに従い時系列で更新されていく系である。シンプル。エレガント。そして恐ろしいほどに——教室そのものだ。
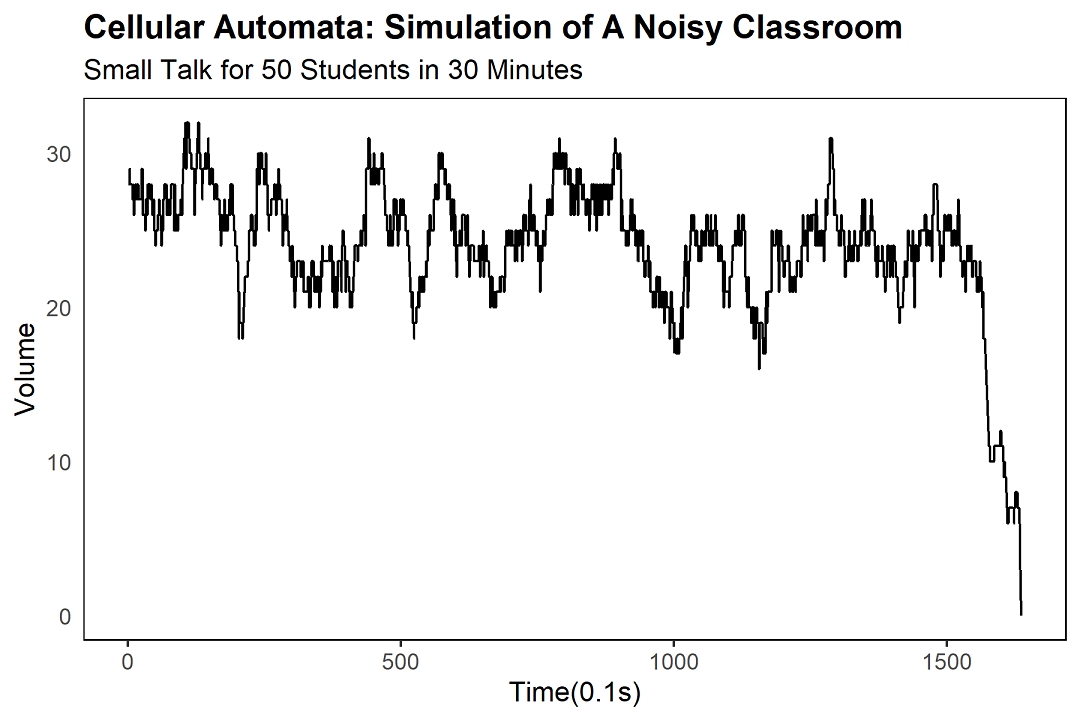
我々のモデルでは、教室は五十個のセルの列として表現される。一セルが一生徒。各生徒は「話す(正の値)」か「聞く(負の値)」の二状態を持つ。値がゼロに達した生徒は、発話を終えた束の間の静寂にあり、次の瞬間には運命のルーレットが回り、-100から+100の新たな値がランダムに割り振られる。
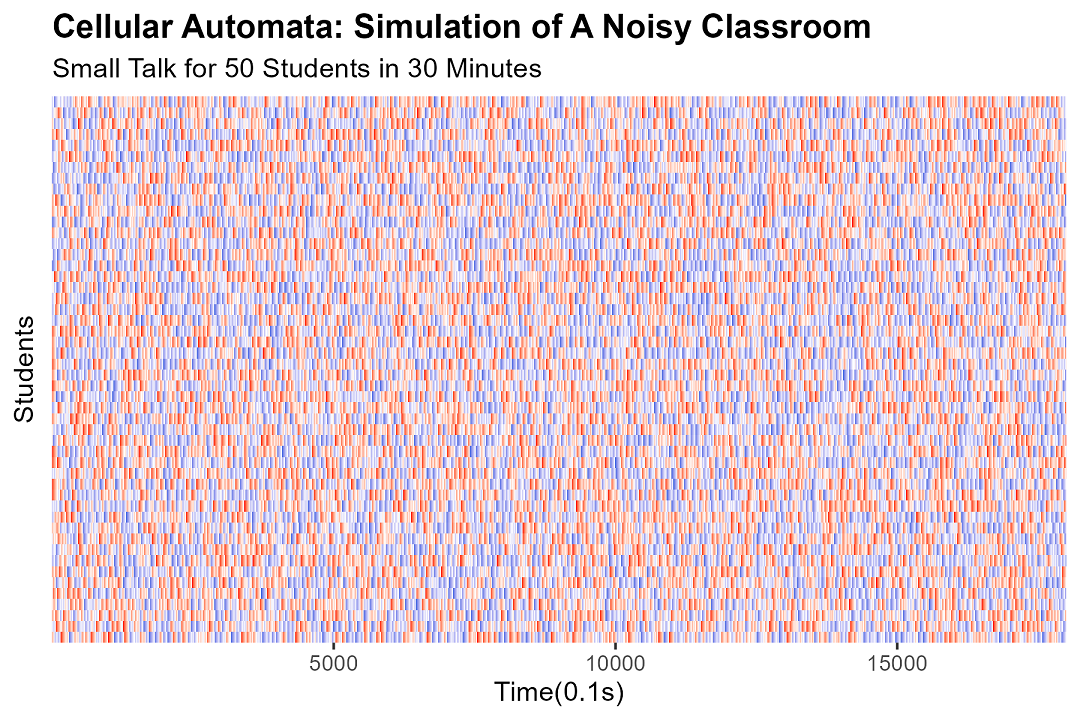
時間ステップを0.1秒に設定し、一回の発話は最大10秒——すなわち100ステップ。+100で開始した生徒は、100ステップ後にゼロに達し、再ランダム化される。数分後の教室は、赤(話す)と青(聞く)が織りなす、壮絶なるカオスのタペストリーだ。
18,000ステップ。すなわち現実時間にして三十分間。これはちょうど、金曜午後の平均的十代の注意力の限界に等しい。俺は知っている。俺が、その十代だったからだ。
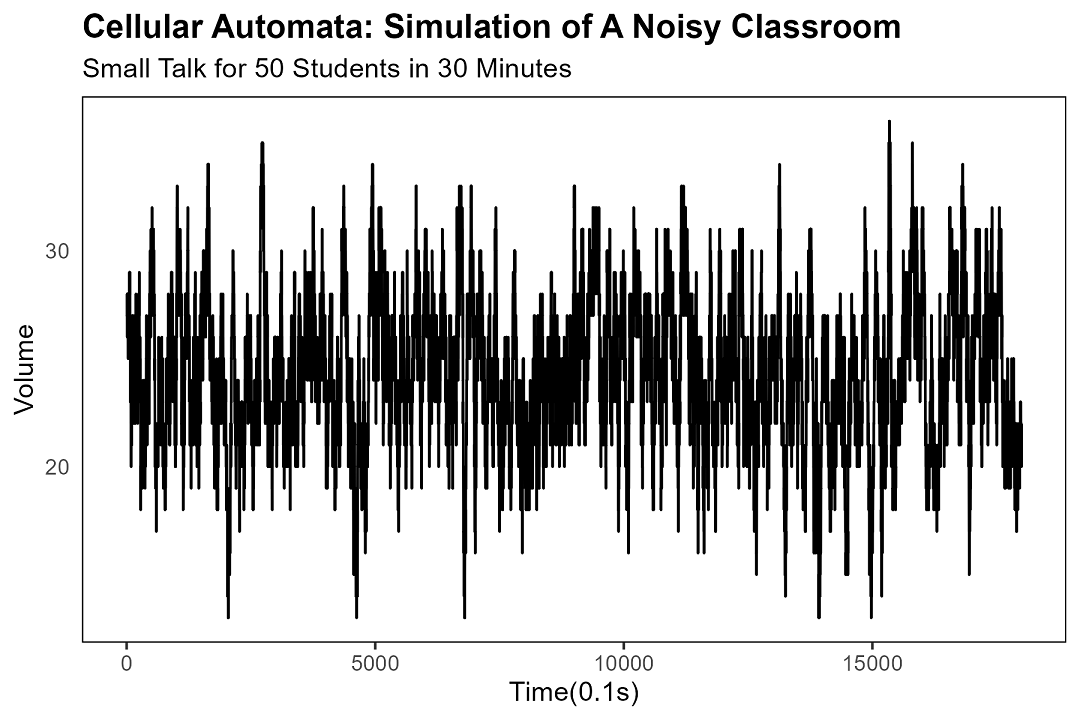
音量の指標として、現時点で発話中の生徒数をカウントする。「厳密な定量分析」が示すのは、ただの「頭数」である。されど——この単純さこそが真理を映す鏡。
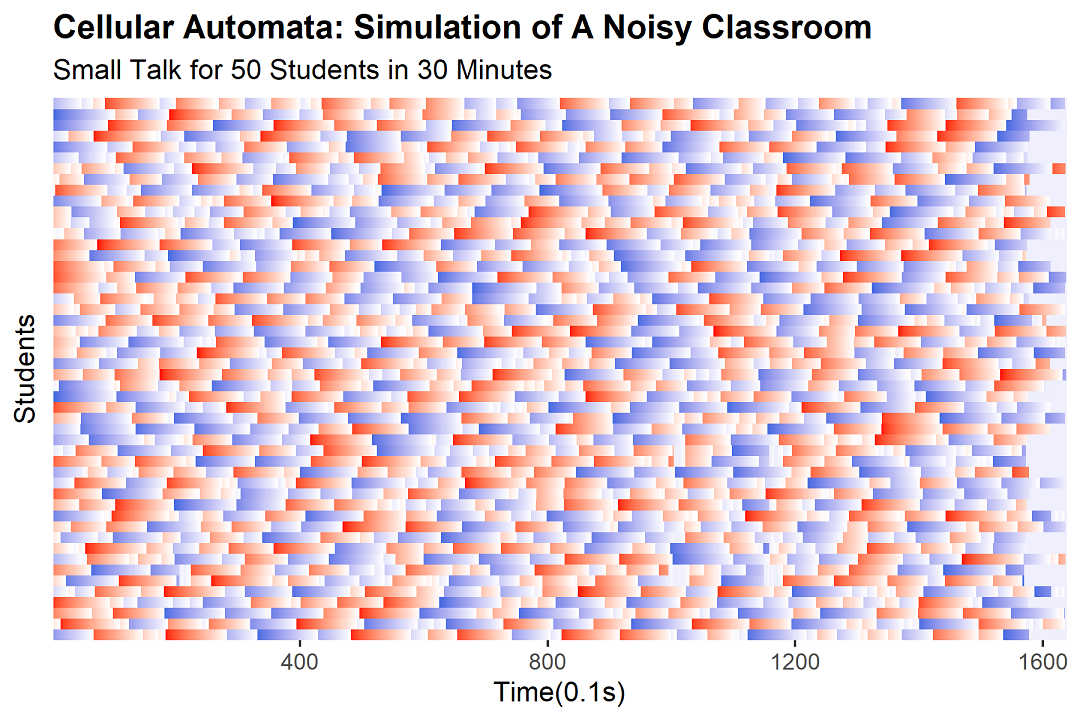
フリーズ反応——そして世界は凍りつく
ここに、モデルに真の深淵をもたらす要素を投入する——各生徒の「心理的影の面積」、すなわち最小凍結音量 $x$ である。
この $x$ は、当該生徒が「異変」を察知し凍結状態に入る閾値を意味する。教室で同時に話している人数が $x$ を下回った瞬間、その生徒の魂は警告を発し、一秒間の「停・看・聴」状態に陥る。十七人以下の話者でパニックを起こす深き影の持ち主もいれば、自分と隣の友だち二人だけになるまで気づかない鈍感なる勇者もいる。この差異こそが、モデルに生命を吹き込む神の一手だ。
実装は単純明快。前ステップの話者数 < $x$ なら、当該生徒の状態を -10 に設定(= 10ステップ = 1秒間の凍結)。そうでなければ、元のルールで推移させる。
そして——凍結は連鎖する。一人が凍れば総話者数が減り、それが次の者の $x$ を下回り、また凍る。この正のフィードバックこそが、「自然発生の沈黙」を引き起こす原動力である。
本シミュレーションにおいて、自然発生の沈黙は約160秒で発現する(現実においては五分で来ることもあれば、五秒で来ることもあり、あるいは永遠に来ぬまま誰かが職員室に呼ばれることもある)。
パラメータの調律——神は細部に宿る
集団的暗黙知が発現するか否かは、システムの規模に非線形に依存する。初期値の微かな揺らぎが、天と地ほどの差異を生む——複雑系の宿命だ。ゆえに、パラメータ設定には細心の注意を要する。
本モデルでは、各セルの初期状態は完全にランダムで、唯一系統的に操作する変数は各生徒の最小凍結音量 $x$ のみ。
現実との接点を保つため、最も鈍感な生徒の $x$ 下限は 3 とする(この男は、教室内に話し手が自分含め四人になるまで何も気づかない豪傑である)。$x$ の上限については、現実から直接導出できないため、シミュレーションによる探索で決定する。
$x_{min}$ = 3、$x_{max}$ = 17 の条件で、一万回の試行を実施。各回で、五十名の $x_i$ の総和 $S_x = \Sigma x_i$ と、自然沈黙の初回出現時間 T を記録した。
| Estimate | Std.Error | t | P | Sig. | |
|---|---|---|---|---|---|
| (Intercept) | 6.0676688 | 0.0829139 | 73.18 | <2e-16 | *** |
| $S_x$ | -0.0056127 | 0.0001652 | -33.97 | <2e-16 | *** |
表1:自然沈黙の発生時間と学生の「心理的影面積」総和の線形回帰
さらに $x_{max}$ を15から19まで段階的に変化させ、各水準で数千回の試行を実施。三十分以内に自然沈黙が生起した頻度からロジスティック回帰モデルを構築した。
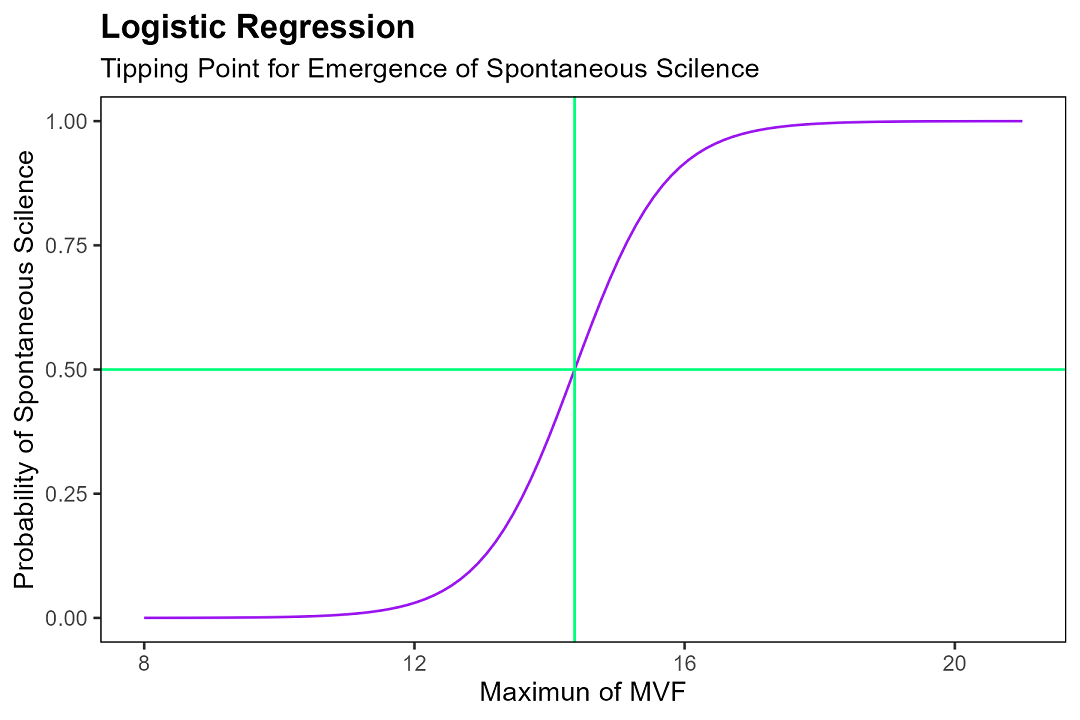
図6:ロジスティック回帰モデル $y = [1 + e^(-1.463x + 21.019)]^{-1}$ と臨界点
この数式が示すところは、要するに——教室の話者数が臨界値を下回った瞬間、氷結の連鎖が誘発され、教室は急速に静寂へと収束する。三十分の自習時間において、五十名中の最大 $x$ が臨界点14を超えるとき、自然沈黙はほぼ不可避となる。19を超えたならば——それはもはや運命だ。確率は天を衝く。
結語
「凍結・逃走・闘争(Freeze, Flight, Fight)」——この三つの F は、動物界において、生命が脅威に直面した際の共通した行動傾向である。科学的知見によれば、捕食者に狙われた被食者にとって「停止」という反応は、しばしば最も合理的な選択となる。これは、肉食哺乳類の視覚野ならびに網膜が、進化の過程で動体の検出に極めて高度に特化してきたことに起因する。すなわち、「停まり、見て、聴く」ことで捕食者の目を逃れた個体こそが、自然淘汰の篩をくぐり抜け、生存と繁殖の機会を次世代に繋いできたのである。我々ヒトの大脳辺縁系に刻まれたこの警戒評価機構は、長大な進化史の中で形成され、現在もなお我々の行動基盤の深層に根差している。
興味深いことに、英語圏にはこのような唐突な静寂を指して「天使が通る(An angel passing by)」と表現する慣用句がある。合理性では説明し難い瞬間の静けさに、人々が詩を見出した——その感性そのものが、人間の文化の豊饒さを物語っているように思われる。
思うに、我々の前に現れた教師たちの中には、まさしく「天使」と呼ぶにふさわしい存在が、確かにいたのではあるまいか。振り返ってみると、人生の最初の十数年のうちに、ほんの一年か二年だけ現れ、そして立ち去っていった人々の中に、純粋にこちらを思い、導こうとしてくれた人々がいたことに、歳を重ねるほどに気づかされる。自律によって良い習慣を身につけることがいかに困難かを知ったとき、かつての厳しい指導が、過酷な競争の中で克服すべき怠惰と先延ばしの習性に抗うための、ささやかながら確かな鎧であったと、ようやく理解できるようになった。
そうした人々は、人生の先へ進むほどに出会うことが少なくなる。だからこそ——どうか、「静かにしなさい」と叱ってくれた天使たちに、遅まきながら感謝の言葉を伝えてほしい。
最後になるが、教室という舞台にはいつも、見せしめとして犠牲となる者が一人は必要だった。遊びも結構、冗談も結構——されど、教師をあまり本気で怒らせるものではない。
付録一:セル・オートマトン実装
i <- 1
timestep <- 1
timeout <- 18000
students <- 50
t0 = round(runif(students, -100, 100))
ca <- cbind.data.frame(t0)
volume <- vector(mode="numeric")
volume[timestep] <- sum(ca[,timestep] > 0)
mentalshadow <- round(runif(students, 3, 19))
while(volume[timestep] != 0){
for(i in 1:students){
if(volume[timestep] < mentalshadow[i]){
ca[i, timestep + 1] <- -10
}
else {
if(ca[i, timestep] == 0) {
ca[i, timestep + 1] <- sample(-100:100, 1)
}
else if(ca[i, timestep] > 0) {
ca[i, timestep + 1] <- ca[i, timestep] - 1
}
else {ca[i, timestep + 1] <- ca[i, timestep] + 1}
}
}
colnames(ca)[timestep+1] <- paste0("t", timestep)
timestep <- timestep + 1
if(timestep == timeout){volume[timestep] = 0}
else{volume[timestep] <- sum(ca[, timestep] > 0)}
}
付録二:ベクトル計算における同期化の罠
はじめに
R言語におけるベクトル演算(ifelse() 等)は、C言語で実装された内部関数により要素ごとの処理を一括で行うため、同等の for ループと比較して著しく高速である。この特性から、シミュレーションデータを規則に基づき生成する際には、ベクトル演算を優先することが一般的な慣行となっている。しかし、ランダムな値の割り当てを含む処理においては、ベクトル演算が意図せずして個体間の独立性を破壊し、非同期的であるべき状態遷移に「lockstep(同期歩調)」パターンを導入する危険性がある。
問題:同期進化がもたらす異常パターン
本論文のシミュレーションでは、各個体(学生)の状態値は以下の二規則に従って更新される:
- 状態が正の場合、1ずつ減少する。状態が負の場合、1ずつ増加する。
- 状態が 0 に達した場合、新たな乱数値が割り当てられる。
すなわち、素朴な実装としては:
ifelse(state == 0, sample(new_values, 1), state ± 1)
このコードは、表面上は与えられた規則を正しく実装しているように見える。しかし、ここで sample(-100:100, 1) はベクトル全体に対して単一の乱数値を生成し、その同一の値を、当該タイムステップにおいて状態が 0 である全ての個体に一括代入する。その結果、同一タイムステップで偶然に状態 0 を迎えた複数の個体は、完全に同一の新状態値を取得し、以降の全タイムステップにわたって完全に同一の進化軌跡を辿ることになる。
set.seed(1)
ca <- data.frame('step0' = round(runif(50, min = -100, max = +100)))
for (t in 1:100) {
ca_rule <- ifelse(ca[, t] == 0, sample(-100:100, 1),
ifelse(ca[, t] > 0, ca[, t] - 1, ca[, t] + 1))
ca[,ncol(ca) + 1] <- ca_rule
colnames(ca)[ncol(ca)] <- paste0("step", t)
}
rownames(ca) <- paste0("Student ", 1:50)
このコードの実行結果として、複数個体が同一の軌跡を描く「lockstep」現象が確認される。
pheatmap(ca,
cluster_rows = TRUE,
cluster_cols = FALSE,
show_rownames = TRUE,
show_colnames = FALSE,
treeheight_row = 0)
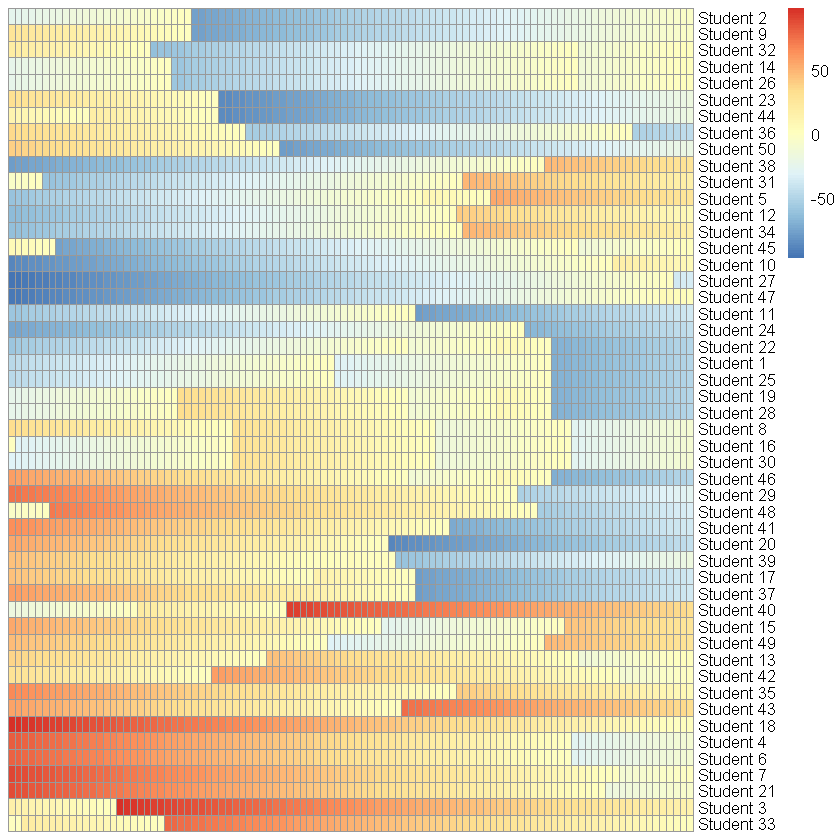
本現象がシミュレーションの妥当性に及ぼす影響は以下の通りである:
- 上図に示されるように、いくつかの個体の状態遷移は完全に同一の軌跡を辿り、事実上区別不可能となる。
- 本来仮定されている個体間の異質性が破壊される。
- 影響を受ける個体の割合が十分に大きい場合、マクロなシミュレーション結果そのものが、意図した期待値から系統的に乖離する可能性がある。
要するに、この実装は個体レベルの独立性および集団レベルの異質性の両方を損なうため、科学的妥当性の観点から看過できない問題を内包している。
解決策:個別乱数割り当てによる独立性の担保
個体間の独立性を維持するには、状態 0 への到達時に、各個体が独立に生成された乱数を取得することを保証しなければならない。実装方針としては、まず当該タイムステップで状態 0 に該当する全個体のインデックスを which() により取得し、その個数分の乱数を sample() により一括生成した上で、該当位置にのみ代入する方法が有効である。これにより、ベクトル演算の高速性を維持しつつ、「lockstep」問題を完全に回避できる。
vectorized_update <- function() {
ca <- data.frame('step0' = round(runif(50, min = -100, max = +100)))
for (t in 1:1000) {
zero_indices <- which(ca[, t] == 0)
new_random_values <- sample(-100:100, length(zero_indices), replace = TRUE)
ca_rule <- ifelse(ca[, t] == 0, NA,
ifelse(ca[, t] > 0, ca[, t] - 1, ca[, t] + 1))
if (length(zero_indices) > 0) {
ca_rule[zero_indices] <- new_random_values
}
ca[, ncol(ca) + 1] <- ca_rule
colnames(ca)[ncol(ca)] <- paste0("step", t)
}
}
比較のため、for ループにより個体ごとに個別の乱数生成を行う実装を以下に示す。本方式では個体間の独立性が保証される一方、ループ回数に比例した計算時間の増大を伴う。
loop_update <- function() {
ca <- data.frame('step0' = round(runif(50, min = -100, max = +100)))
for (t in 1:1000) {
ca_rule <- numeric(50)
for (i in 1:50) {
if (ca[i, t] == 0) {
ca_rule[i] <- sample(-100:100, 1)
} else if (ca[i, t] > 0) {
ca_rule[i] <- ca[i, t] - 1
} else {
ca_rule[i] <- ca[i, t] + 1
}
}
ca[, ncol(ca) + 1] <- ca_rule
colnames(ca)[ncol(ca)] <- paste0("step", t)
}
}
性能比較
ベクトル化による改良版(vectorized_update)とループ版(loop_update)の実行時間を system.time() により測定した結果を以下に示す。測定は各5回実施した。
replicate(5, system.time(vectorized_update())["elapsed"])
0.340000000000146
0.319999999999709
0.340000000000146
0.270000000000437
0.25
replicate(5, system.time(loop_update())["elapsed"])
2.5
2.42000000000189
2.40999999999985
2.90000000000146
2.88999999999942
ベクトル化版の平均実行時間は約 0.30 秒、ループ版は約 2.62 秒であり、ベクトル化版は約 8.7 倍の速度向上を達成している。この性能差の主要因は、(1) ifelse() の内部実装が C 言語により最適化されていること、(2) which() による一括インデックス検索の効率性、(3) sample() の呼び出し回数がループ版(個体数 × タイムステップ)に対してベクトル化版ではタイムステップ数に比例すること、の三点に集約される。
結論
ベクトル演算は R 言語における高速なデータ処理の中核を成す技法であり、大規模データセットや長時間シミュレーションにおいては不可欠の最適化手段である。しかしながら、ランダム値の割り当てを含む処理では、全要素に同一の乱数が適用されるという特性が、個体ベース・シミュレーションの根幹である「個体間の独立性」を損なう危険性を孕む。この問題は、which() による条件該当インデックスの事前抽出と、該当個体数に応じた乱数の一括生成・個別代入によって回避できる。これにより、ベクトル演算の計算効率を維持しつつ、シミュレーションの科学的妥当性を担保することが可能である。