要旨
本研究は、海水(SEA)と淡水(TAP)の環境下で飼育したニホンウナギの三つの重要組織(脳、生殖腺、胸鰭)について差次的発現遺伝子(DEG)解析を行い、塩分適応過程におけるトランスクリプトーム応答機構の解明を目的とする。
主な知見:
- 包括的分析(組織効果の制御)により422個のDEG(発現上昇303、発現低下119)を同定した
- 脳は塩分変化に最も敏感で、計500個のDEGが検出された
- 三組織すべてで共通して有意な応答を示した中核的遺伝子は8個であり、いずれも海水環境下で発現上昇していた
主な結論:
- ニホンウナギは塩分変化に対して組織特異的なトランスクリプトーム応答を示し、脳の応答が最も強く、胸鰭が最も保守的であった。
- 海水環境はより多くの遺伝子を活性化し(包括的分析:72%が発現上昇)、海水の塩分環境が特定の遺伝子発現プログラムを促進することを示唆する。
- 組織ごとの応答パターンの差異は、組織特異的な機能が環境塩分に対してそれぞれ異なる適応戦略をとることを反映している。
一、材料と方法
1.1 試料情報
実験には、2017年5月31日に泉州地域で購入した非野生の養殖ウナギを用いた。購入時に、体重300g〜1000gで、体形がふっくらとし、胸鰭が丸みを帯びて色が濃く、背部が黒色で腹部が銀白色の個体を選別した。これらは降海前の雌の銀ウナギの典型的な特徴であり、形態学的な雌雄判別の正確度は約90%である1。
5尾の非野生ウナギからランダムに3尾を選び塩分馴致を行い、淡水飼育のウナギと対照群を形成した。毎日定時に6.6‰の海水塩を添加し、5日目には実験群のウナギの環境塩分濃度が約30‰(海水の塩分は約35‰)に達し、ニホンウナギの降海回遊時の塩分変化過程を模擬した。
サンプリングに際しては、全5尾のニホンウナギを氷水処理により麻痺させた後、形態指標の測定および解剖を行った。各ニホンウナギから脳、生殖腺および胸鰭をRNAシーケンシング用試料として採取し、RNAの加水分解を防ぐため、採取後直ちに液体窒素中で凍結保存した。
| 試料 | 体長(mm) | 体重(g) | 水平眼径(mm) | 垂直眼径(mm) | 眼間隔(mm) | 吻長(mm) | 胸鰭長(mm) | 胸鰭幅(mm) | 肛門-腹鰭起始端間(mm) |
|---|---|---|---|---|---|---|---|---|---|
| SEA_1 | 695 | 553 | 12 | 11 | 15 | 12 | 35 | 16 | 5 |
| SEA_2 | 650 | 570 | 10 | 9 | 13 | 13 | 30 | 20 | 4 |
| SEA_3 | 680 | 600 | 11 | 11 | 13 | 13 | 30 | 16 | 4 |
| TAP_1 | 675 | 610 | / | / | / | / | 30 | 20 | 8 |
| TAP_2 | 715 | 650 | 9 | 10 | 15 | 16 | 27 | 18 | 5 |
注:
- SEA_N:実験群、TAP_N:対照群;
- 体長:吻端から尾鰭基部までの長さ;
- 水平眼径:眼窩前縁から後縁までの幅;垂直眼径:眼窩上縁から下縁までの幅;
- 眼間隔:両眼中心間の幅;
- 吻長:眼窩前縁から吻先端までの長さ;
- 胸鰭長:胸鰭基部から末端までの長さ。
本解析は合計15試料を含み、3組織および2塩分処理条件を網羅する:
| 組織 | 海水群(SEA) | 淡水群(TAP) |
|---|---|---|
| 脳(brain) | BR_SEA_1, BR_SEA_2, BR_SEA_3 | BR_TAP_1, BR_TAP_2 |
| 生殖腺(gonad) | GD_SEA_1, GD_SEA_2, GD_SEA_3 | GD_TAP_1, GD_TAP_2 |
| 胸鰭(pectoral fins) | PF_SEA_1, PF_SEA_2, PF_SEA_3 | PF_TAP_1, PF_TAP_2 |
1.2 データ前処理
本解析はトランスクリプトームシーケンシングデータに基づき、参照配列を用いないTrinityアセンブリにより16,920個の推定遺伝子を得た。シーケンシング深度が解析結果に与える影響を低減するため、遺伝子発現量に対するフィルタリングを行い、少なくとも3試料においてカウント値が10未満の遺伝子を除去し、最終的に16,866個の遺伝子を以降の差次的発現解析に供した。
1.3 統計解析手法
本解析では二種類の実験デザイン戦略を採用した(有意性閾値:padj < 0.05):
| 特徴 | 包括的分析(組織効果の制御) | 各組織別分析 |
|---|---|---|
| デザイン式 | ~tissue + treatment | ~treatment |
| 試料範囲 | 全15試料 | 各組織5試料 |
| メタデータ | tissue および treatment を含む | treatment のみを含む |
| 目的 | 組織を横断して一貫した処理効果の検出 | 特定組織内における処理効果の検出 |
包括的分析(多因子モデル)
design = ~tissue + treatment
包括的分析では多因子線形モデルを用い、組織間のベースライン差異を制御した上で、組織を横断して一貫した処理効果を検出する:
$$ log2(μ_ij) = β_0 + β_{tissue_j} + β_{treatment} · X_{treatment} $$ここで:
- $β_0$:切片(ベースライン発現水準)
- $β_{tissue_j}$:組織効果 — 異なる組織(脳、生殖腺、胸鰭)間のベースライン発現差異
- $β_{treatment}$:処理効果 — 組織差を制御した後の、SEA対TAPの発現変化
組織を共変量としてモデルに組み込むことは、以下を意味する:
- モデルはまず、異なる組織自体の基礎的な発現差異を「差し引く」
- その後、全組織に適用される統一的なtreatment効果を推定する
- 処理効果が異なる組織間で同一であると仮定する
各組織別分析(単因子モデル)
design = ~treatment
各組織内で独立に海水対淡水の差次的発現を検出する。
Wald検定
Wald検定は、モデル中の特定の係数(例えば処理条件の効果)が有意に非ゼロであるかを評価するために用いられる。RNA-seqの差次的発現解析で一般的な負の二項分布モデルでは、各遺伝子に対して次のモデルをあてはめる:
$$ \log(\mu_{ij}) = \beta_0 + \beta_1 X_{condition} + \beta_2 X_{tissue} + \dots $$Wald検定は$\beta_1$(処理条件効果)の有意性を検定するために用いられ、試料数が少ないがモデル適合が比較的安定している状況に適している。
Benjamini-Hochberg(BH)補正
RNA-seqデータは通常、数千から数万の遺伝子を含み、各遺伝子に対してp値が算出される。生のp値をそのまま用いると多数の偽陽性(第一種の過誤)が生じる。BH法は偽発見率(FDR)、すなわち有意と判定された全遺伝子のうち、誤って有意とされるものの期待割合を制御する。多重比較問題(例:20,000遺伝子)において、FDR制御はBonferroni補正よりも緩やかで合理的であり、より多くの潜在的生物学的シグナルを保持できる。BH補正はp値の分布に対して特別な仮定を置かないため、個々の遺伝子の試料数が少ない場合でも、多数の遺伝子の多重検定問題の処理に適している。
二、解析結果
2.1 差次的発現遺伝子の概要
包括的分析の結果(組織効果の制御)
| 指標 | 数値 |
|---|---|
| 総DEG数 | 422 |
| 発現上昇遺伝子(SEA > TAP) | 303 |
| 発現低下遺伝子(SEA < TAP) | 119 |
| 上昇/低下比 | 2.55 |
包括的分析(組織効果の制御)の結果、16,866個の有効遺伝子から422個の差次的発現遺伝子が同定され、そのうち303遺伝子が海水環境下で発現上昇、119遺伝子が発現低下を示し、発現上昇遺伝子の割合は72%に達した。この結果は、ニホンウナギの海水塩分環境への応答が、主として遺伝子発現の活性化として現れることを示している。上昇/低下比が2.55に達したことは、海水環境がより積極的な遺伝子発現プログラムを誘起したことを示唆しており、浸透圧調節、イオン輸送、エネルギー代謝などの適応機構と密接に関連している可能性がある。
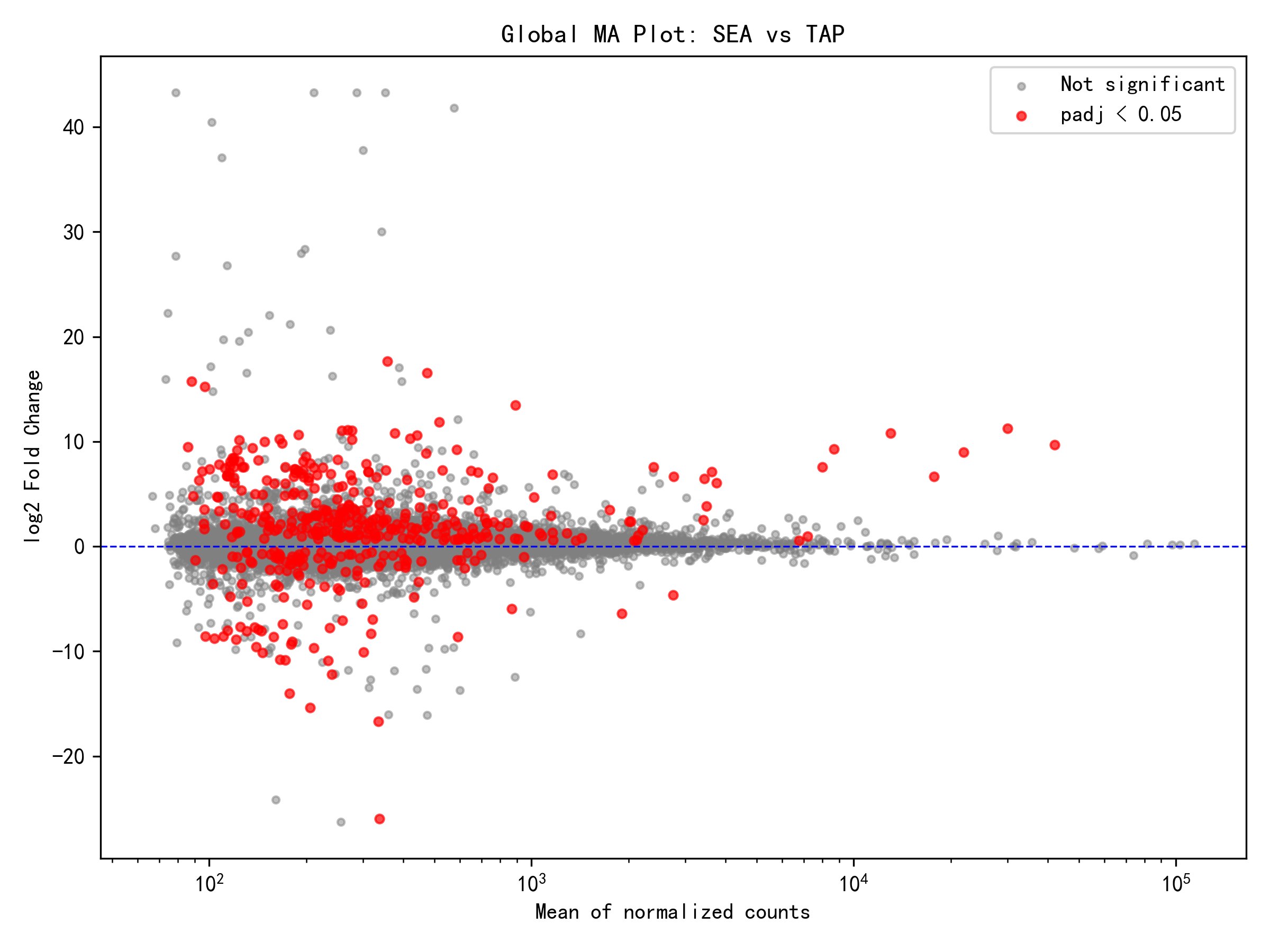
各組織別分析の結果
| 組織 | 総DEG数 | 発現上昇 | 発現低下 | 上昇/低下比 |
|---|---|---|---|---|
| 脳(brain) | 500 | 340 | 160 | 2.13 |
| 生殖腺(gonad) | 131 | 75 | 56 | 1.34 |
| 胸鰭(pectoral_fins) | 97 | 82 | 15 | 5.47 |
各組織別分析では、三組織が顕著に異なる応答パターンを示した。脳組織では最多の差次的発現遺伝子(500個)が検出され、うち340個が発現上昇、160個が発現低下、上昇/低下比は2.13であり、脳が塩分変化に最も敏感であることを示している。これは、神経内分泌による浸透圧適応応答の中枢的調節機能と合致する。生殖腺組織では131個の差次的発現遺伝子が検出され、発現上昇75個、発現低下56個、上昇/低下比は1.34であり、三組織中で最も1:1に近い比率を示した。このことは、生殖腺の塩分変化への応答が比較的バランスのとれたものであり、代謝と生殖機能の協調的調整に関与している可能性を示唆する。胸鰭組織では差次的発現遺伝子数が最も少なかった(97個)が、上昇/低下比は5.47と極めて高く(発現上昇82個、発現低下15個)、強い一方向的応答特性を示した。これは、胸鰭において少数の遺伝子が海水環境により強力に活性化され、直接的な浸透圧調節に関わる機能タンパク質をコードしている可能性を示唆する。
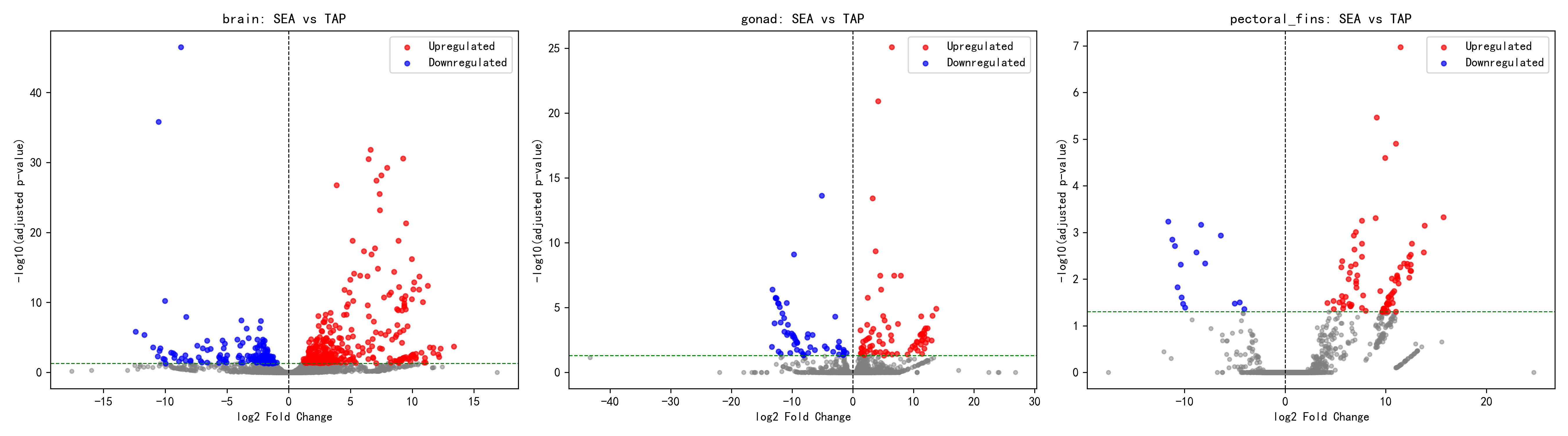
2.2 組織特異的応答の解析
脳(brain)——最も敏感な塩分応答組織
脳は最も強い塩分応答を示し、DEG数(500)は他の組織を大きく上回った。この知見は以下を示唆する:
- 神経調節機構が塩分適応において中核的役割を果たしている可能性がある
- 脳は浸透圧受容器と神経内分泌経路の調節を通じて、全身の塩分適応応答を統御している可能性がある
- 発現上昇遺伝子の高比率(340/500 = 68%)は、海水環境が脳における広範な遺伝子発現プログラムを活性化したことを示す
生殖腺(gonad)——中等度の塩分応答
生殖腺のDEG数(131)は脳と胸鰭の中間に位置し、上昇/低下比(1.34)も1:1に近い。これは以下を反映している可能性がある:
- 繁殖器官が環境塩分の変化に対して一定程度の応答を示す
- 海水環境が生殖腺の代謝および成熟過程に影響を与える可能性がある
- 性別に依存した繁殖戦略が、塩分に対する適応的調節を伴う可能性がある
胸鰭(pectoral_fins)——保守的な塩分応答
胸鰭のDEG数は最も少ない(97)が、上昇/低下比は最も高い(5.47)。このことは以下を示す:
- 胸鰭の塩分変化に対する転写応答は比較的保守的である
- 少数の遺伝子が海水環境下で強力に活性化される
- これらの遺伝子は塩分適応に直接関連するタンパク質(イオンチャネル、浸透圧調節タンパク質など)をコードしている可能性がある
2.3 中核的応答遺伝子(三組織共通有意)
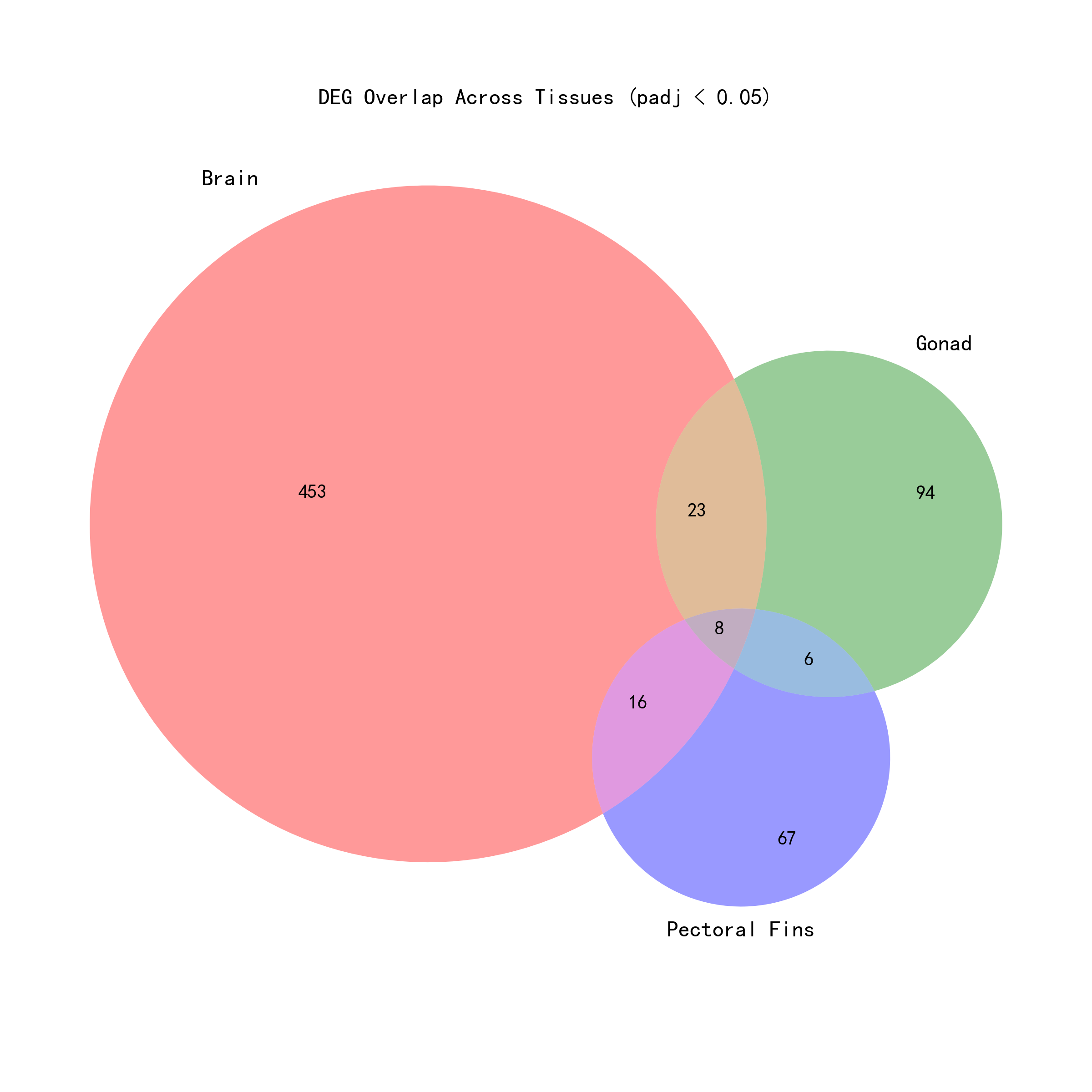
三組織の各組織別分析結果の積集合を取ることにより、8個の中核的差次的発現遺伝子が同定された——これらの遺伝子は三組織すべてにおいて有意な塩分応答を示した(padj < 0.05):
| 遺伝子ID | 脳 log2FC | 生殖腺 log2FC | 胸鰭 log2FC | 平均 log2FC |
|---|---|---|---|---|
| _TRINITY_DN215578_c8_g2_i4 | 11.41 | 10.20 | 11.25 | 10.95 |
| _TRINITY_DN212746_c1_g3_i1 | 8.36 | 11.78 | 9.63 | 9.92 |
| _TRINITY_DN196393_c2_g1_i4 | 7.71 | 4.68 | 12.12 | 8.17 |
| _TRINITY_DN210877_c3_g1_i17 | 8.97 | 4.37 | 11.16 | 8.17 |
| _TRINITY_DN192233_c0_g2_i2 | 7.38 | 4.51 | 12.58 | 8.16 |
| _TRINITY_DN193247_c3_g1_i5 | 9.49 | 3.52 | 7.74 | 6.92 |
| _TRINITY_DN220510_c3_g1_i1 | 9.28 | 4.17 | 5.66 | 6.37 |
| _TRINITY_DN192231_c4_g1_i1 | 5.60 | 5.45 | 6.01 | 5.69 |
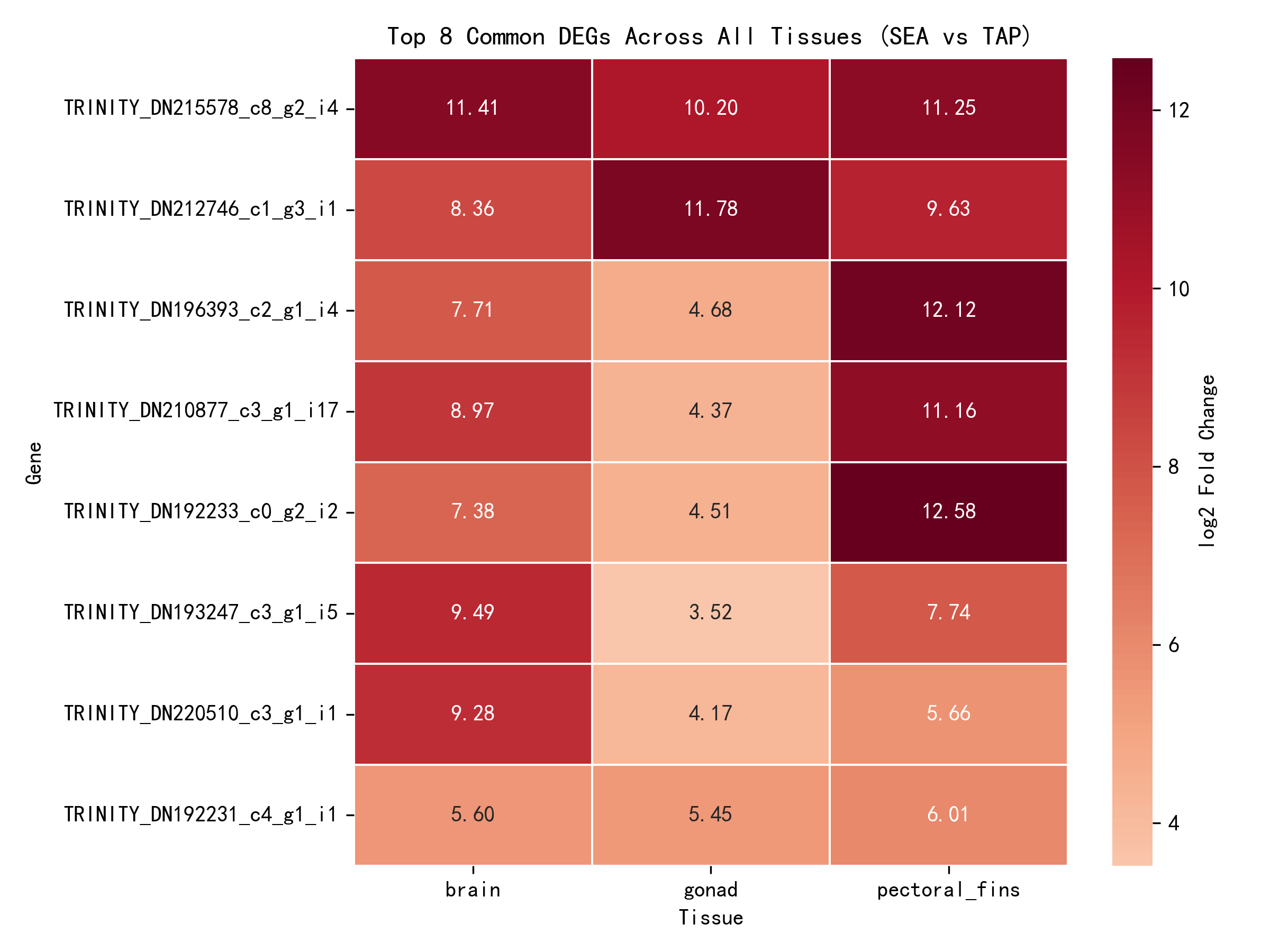
8個すべての中核的応答遺伝子が発現上昇(log2FC > 0)を示し、これらの遺伝子が海水環境下で発現が増強され、ニホンウナギの塩分適応の中核的機構に関与している可能性を示唆する。
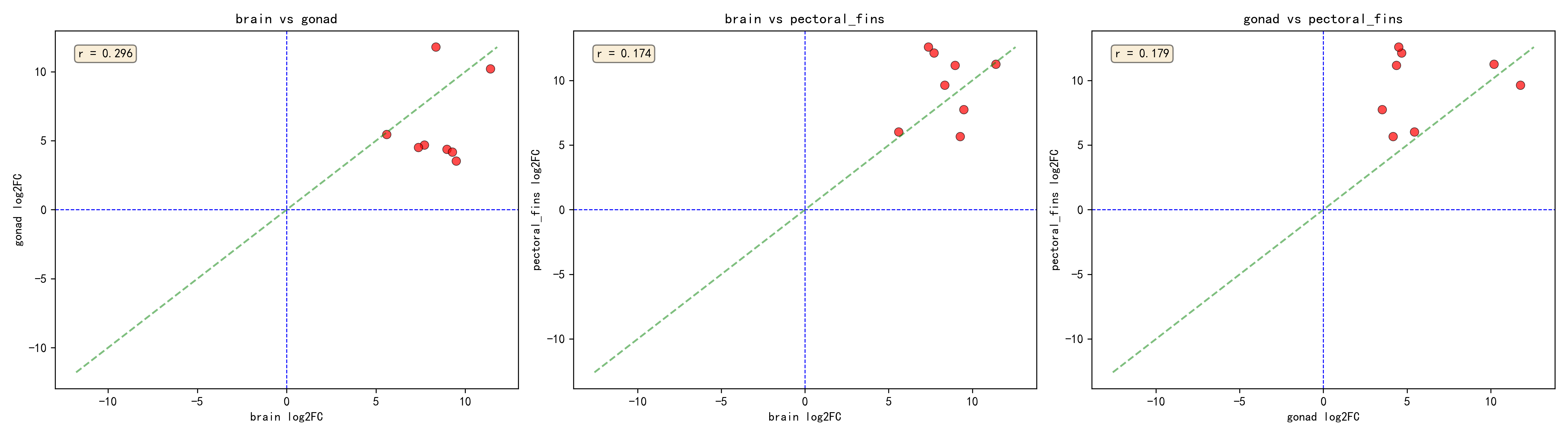
2.4 包括的分析と各組織別分析の交差検証
二つの解析戦略の関係を包括的に理解するため、解析結果に対して交差検証統計を行った:
| カテゴリ | 遺伝子数 |
|---|---|
| 包括的に有意 | 422 |
| 三組織すべてで有意(脳+生殖腺+胸鰭) | 8 |
| 三組織すべてで有意だが、包括的には非有意 | 0 |
| 包括的に有意だが、三組織すべてでは非有意 | 414 |
包括的に有意だが三組織すべてでは非有意な414遺伝子のさらなる分類結果は以下の通り:
| 有意組織数 | 遺伝子数 | 詳細 |
|---|---|---|
| 0組織で有意 | 226 | 各組織別分析ではいずれも非有意 |
| 1組織で有意 | 148 | 104(脳のみ)、31(生殖腺のみ)、13(胸鰭のみ) |
| 2組織で有意 | 40 | 二組織で有意だが、三組織すべてではない |
8個の三組織共通有意遺伝子は、包括的分析でもすべて有意であった。包括的に有意だが三組織すべてでは非有意なケースが大量に存在する——計414遺伝子で、包括的に有意な遺伝子の98%(414/422)を占める。特に注目すべきは、各組織別分析ではいずれも非有意だが包括的分析では有意な226遺伝子である——これらの遺伝子は、包括的分析が組織横断的な統合によって統計的検出力を向上させ、単独組織では効果が弱いが方向の一貫した遺伝子を検出できた可能性がある。
三、結論と考察
本研究は、包括的分析と各組織別分析を組み合わせた戦略により、海水と淡水環境下におけるニホンウナギの三つの重要組織の差次的発現遺伝子を系統的に解析した。包括的分析では組織効果を制御して422個のDEGを同定し、うち303個が発現上昇、119個が発現低下を示した。各組織別分析では、脳の応答が最も強く(500 DEG)、生殖腺がこれに次ぎ(131 DEG)、胸鰭が最も保守的(97 DEG)であった。交差検証により、三組織すべてで有意に発現上昇する8個の中核的応答遺伝子と、包括的分析でのみ有意で各組織別分析では非有意な226個の潜在的応答遺伝子が明らかになった。
包括的分析は組織を共変量としてモデルに組み込み、組織間のベースライン差異を制御した上で組織横断的に一貫した処理効果を検出するものであり、より大きな試料数(n=15)を活用して高い統計的検出力を得る。一方、各組織別分析は各組織内で独立に検定を行い、組織ごとに異なる応答パターンを許容し、三回の独立した補正と積集合によりより厳格な結果を得る。二つの解析戦略の交差検証により、414遺伝子(包括的に有意な遺伝子の98%)が包括的に有意だが三組織すべてでは非有意であることが明らかになった。このうち、226遺伝子は各組織別分析ではいずれも非有意で、包括的分析でのみ検出された。これらの遺伝子の処理効果は方向が一貫しているが、単独組織では効果量が弱く、包括的分析が組織横断的統合により統計的検出力を向上させたものである。148遺伝子は単一組織でのみ有意であり(脳104個、生殖腺31個、胸鰭13個)、40遺伝子は二組織で有意であった。典型的なケースとして、ある遺伝子が脳組織では極めて強い変化を示す(例:log2FC=4)が、生殖腺と胸鰭では変化が弱く(例:log2FC=0.5)、包括的モデルでは大きな試料数を活用して有意な効果を検出できるが、各組織別分析では試料数が少ない(n=5)ために非有意となる場合がある。
三組織すべておよび包括的分析で一貫して有意な8個の中核的応答遺伝子は、そのlog2FCが全組織で5を超えており、これらの遺伝子の塩分変化に対する応答が極めて強いことを示している。注目すべきは、本研究では三組織すべてで有意だが包括的には非有意なケースが認められなかったことである。このようなケースは稀ではあるが起こりうる。例えば、三組織で効果の方向が反対(脳で上昇、生殖腺で低下、胸鰭で上昇)で、包括的モデルではこれらの効果が相互に相殺される場合である。このことは、ニホンウナギの塩分適応における組織特異的応答が、主として効果量の差異として現れ、方向の対立ではないことを示している。8個の中核的応答遺伝子の処理効果は方向が一貫しているだけでなく、各組織における効果がいずれも十分に強く、二つの解析戦略の両方で同時に検出可能であり、最も頑健な塩分応答遺伝子を代表するものとして、さらなる機能研究の価値がある。
各組織別分析で有意な遺伝子のうち、脳に特有の有意遺伝子数が最も多く(104個)、かつ脳は包括的に有意だが三組織すべてでは非有意な414遺伝子の大部分を占めている。このことは、脳が浸透圧受容器および神経内分泌調節中枢として機能することと合致し、脳が塩分感知と全身的適応応答の調節において主導的役割を果たすことを示唆する。包括的分析は、単独組織では効果が弱いが方向の一貫した遺伝子を検出することができ、これら226個の潜在的応答遺伝子は、普遍的に存在するイオンチャネルや浸透圧応答エレメントなど、基礎的な細胞浸透圧調節機構に関与している可能性があり、塩分適応に不可欠な基盤的調節ネットワークを構成していると考えられる。
塩分適応過程の後、ニホンウナギの生殖腺組織において、脂肪代謝関連遺伝子の変化が観察された。TRINITY_DN187689_c0_g1_i9の遺伝子機能アノテーション結果から、この推定遺伝子の後半部分はアジアアロワナ(Scleropages formosus)のPlin2(Perilipin 2)遺伝子の複数のtranscript variant(転写スプライシングバリアント)と類似した塩基配列を有し、RNA発現量が増加していた(p値 = 0.0027、ただし補正後は非有意 padj = 0.2023)。TRINITY_DN192327_c4_g3_i14はアジアアロワナのスフィンゴミエリン合成酵素1(Sphingomyelin synthase 1、Sgms1)に類似した遺伝子であり、降海後にそのRNA発現量が低下した。
Perilipin 2は脂肪分化関連タンパク質であり、当初ADRPと命名され2、その転写レベルの上昇は脂肪細胞分化初期の兆候の一つであるが、決定的なものではない。Plin2遺伝子をノックアウトしても脂肪分化過程は正常に進行しうる3。確かなのは、Plin2がスフィンゴミエリン(SM)およびホスファチジルコリン(PC)と高い親和性を持つことである4。しかし、その機能は未だ不明であり、脂質化されたタンパク質に保護バリアを提供し、脂肪沈着を促進し、トリグリセリドの代謝回転を低下させるものと考えられている5'6。
脂肪の合成、分化および沈着はニホンウナギの生殖腺発達と不可分の関係にある。張潔明ほかによるニホンウナギ卵巣発達のステージ分類と記述によれば、発達適応期において、ニホンウナギの卵母細胞に一層または多層の脂肪胞が出現し始め、発達始動期に入ると脂肪胞の厚さはさらに増加して配列がより密になり、成長期に厚さが最大に達した後、減少・疎鬆化していく7。ニホンウナギは降海回遊を開始した後、一切の摂餌を停止し、身体エネルギーと生殖腺のさらなる発達は脂肪の利用に依存し、筋肉組織中の脂肪動員が卵巣発達の物質的基盤となる8。筋肉組織のリン脂質代謝には一般に三つの異なる経路があり、(1)リソソームから放出されるホスホリパーゼによる消化、(2)スフィンゴミエリナーゼによる加水分解でスフィンゴミエリンを生成、(3)ホスファチジルコリンの合成、が含まれる9。我々は、降海過程においてニホンウナギのPlin2 RNA発現量が上昇し、Sgms1が抑制されるのは、脂肪が動員されて卵巣に到達した後、ホスホリパーゼによる消化もスフィンゴミエリン合成も行われず、発達適応期に沈着を開始し、魚卵に必要な大量のホスファチジルコリンの準備を提供するためではないかと推察する。
四、研究の限界と展望
本研究には以下の限界がある。第一に、試料数が少なく(SEA群3個体、TAP群2個体)、統計的検出力が制限され、効果量が中程度の遺伝子の一部が検出できなかった可能性がある。第二に、実験では段階的に上昇させる塩分馴致方式を採用したが、これはニホンウナギの自然の降海過程で経験する塩分変化とは差異がある可能性がある。第三に、本研究はトランスクリプトームレベルの解析であり、今後はqPCR、ウェスタンブロットなどの実験により重要遺伝子の発現変化を検証し、機能実験(遺伝子ノックアウト/過剰発現など)を通じて、塩分適応における具体的な役割を解明する必要がある。
また、本研究で同定された8個の中核的応答遺伝子については、系統的な機能アノテーション解析を完了している。これらの遺伝子は海水と淡水環境下で高度に一貫した発現応答パターンを示し、ニホンウナギの塩分適応の中核的調節経路に関与する可能性を示唆しており、重要な生物学的研究価値を持つ。しかしながら、これらの遺伝子の調節ネットワーク解析作業はなお進行中であり、関連成果は未整理かつ未公表であるため、本論文では機能アノテーションの予備的結果のみを提示する。これらの中核的応答遺伝子がどのように協調して浸透圧バランス、イオン輸送およびエネルギー代謝などの重要な生理過程を調節するかについては、今後さらにタンパク質相互作用オミクス、メタボロミクスおよび機能検証実験を組み合わせて詳細に解明する必要がある。後続研究では、これらの遺伝子の塩分適応過程における上流-下流の調節関係の解析に焦点を当て、ニホンウナギの塩分応答の完全な調節ネットワークを構築し、ウナギの回遊の分子機構の理解に理論的基盤を提供することを目指す。
郭弘艺, 魏凯, 谢正丽, ほか. 长江口银色鳗的形态指标体系及其雌雄鉴别[J]. 水产学报, 2011, 35(1): 1-9. ↩︎
BRASAEMLE D L, BARBER T, WOLINS N E, et al. Adipose differentiation-related protein is an ubiquitously expressed lipid storage droplet-associated protein[J]. Journal of Lipid Research, 1997, 38(11):2249-2263. ↩︎
CHANG H J, LAN L, PAUL A, et al. Protection against Fatty Liver but Normal Adipogenesis in Mice Lacking Adipose Differentiation-Related Protein[J]. Molecular & Cellular Biology, 2006, 26(3):1063-1076. ↩︎
MCINTOSH A L, STOREY S M, ATSHAVES B P. Intracellular lipid droplets contain dynamic pools of sphingomyelin: ADRP binds phospholipids with high affinity[J]. Lipids, 2010, 45(6):465-477. ↩︎
LISTENBERGER L L, OSTERMEYER-FAY A G, GOLDBERG E B, et al. Adipocyte differentiation-related protein reduces the lipid droplet association of adipose triglyceride lipase and slows triacylglycerol turnover[J]. Journal of Lipid Research, 2007, 14(12):2751-2761. ↩︎
STRACKE F, MA H, THIEL E. Singlet molecular oxygen photosensitized by Rhodamine dyes: correlation with photophysical properties of the sensitizers[J]. Journal of Photochemistry & Photobiology A Chemistry, 1999, 126(1–3):51-58. ↩︎
张洁明, 柳凌, 郭峰, ほか. 人工诱导日本鳗鲡性腺发育组织学研究[J]. 中国水产科学, 2007, 14(4): 593-601. ↩︎
张利红, 张为民, 林浩然, ほか. 性类固醇激素诱导雌性日本鳗鲡性腺发育过程中钙和脂肪的动员[J]. 中山大学学报自然科学版, 2002, 40(2):86-88. ↩︎
SPENCE M W, CLARKE J T, COOK H W. Pathways of sphingomyelin metabolism in cultured fibroblasts from normal and sphingomyelin lipidosis subjects[J]. Journal of Biological Chemistry, 1983, 258(14):8595. ↩︎