读到一篇有意思的论文。Greig 和 Travisano 在 Biology Letters 上发了篇文章,研究酵母的"作弊"行为。
酵母会分泌 invertase 到细胞外消化蔗糖,消化的糖大家都可以用——这就有意思了。一个细胞可以选择"作弊":偷用邻居分泌的酶,自己却不分泌。研究者把有功能 SUC2 基因的酵母叫"合作者",把删除了 SUC2 基因的叫"作弊者",然后把它们放在一起竞争。
结果很反直觉:
- 在稀疏的群体里(低社交密度),作弊者的 fitness 只有 0.87——比合作者差
- 在密集的群体里(高社交密度),作弊者的 fitness 高达 1.19——比合作者强
为什么?社交密度越高,合作者越容易遇到其他合作者。大家一起分泌酶,公共池塘变大,每个人的收益都高。但这时候作弊者混进来,单方面享受公共成果还不付成本,收益爆炸。
当密度极高时,作弊者几乎总能找到合作目标,偷到的比自己分泌的还多。合作者反而被拖累。
这完美符合我的直觉。但我想自己做一遍——不是验证论文结论,而是想亲手"看见"这个过程,把方程写进格子,看数字跑起来。
模型设定
$n \times n$ 的网格,population density 控制每格放置 agent 的概率。Agent 有两种策略:C(合作,分泌酶)和 D(作弊,不分泌)。
两两相遇时玩标准囚徒困境,payoff 矩阵:
$$ \begin{pmatrix} R=3 & S=0 \\ T=5 & P=1 \end{pmatrix} $$R 是合作-合作的奖励,T 是背叛的诱惑,S 是被背叛者的收益,P 是双双背叛的惩罚。按经典设定:$T > R > P > S$,且 $2R > T + S$(互惠合作优于反复背叛)。
每一步:
- Agent 在 Von Neumann 邻域(上下左右四格)找邻居
- 两人玩一把博弈
- 比较这一步的得分 $\pi$
- 以正比于收益差的概率模仿邻居策略
只看当期得分。不记历史,不做规划。
理论预测
复制子动态(Replicator Dynamics)给出了平均场下的方程:
$$\frac{dx}{dt} = x(1-x)[\pi_C - \pi_D]$$$x$ 是合作者比例,$\pi_C$ 和 $\pi_D$ 是两种策略的期望单步收益。
收益依赖于相遇概率。在平均场近似下,合作者遇到合作者的概率是 $x$,遇到作弊者是 $1-x$;作弊者反之:
$$\pi_C = x \cdot R + (1-x) \cdot S$$$$\pi_D = x \cdot T + (1-x) \cdot P$$
代入:
$$\pi_C - \pi_D = x(R - T) + (1-x)(S - P)$$用具体数值 $R=3, S=0, T=5, P=1$:
$$\pi_C - \pi_D = x(3-5) + (1-x)(0-1) = -2x -1 + x = -1 - x$$$\pi_C - \pi_D < 0$ 恒成立——无论 $x$ 多大,合作者的当期收益永远低于作弊者。
这意味着 $\frac{dx}{dt} = x(1-x)(-1-x) < 0$。合作者比例只能单向衰减,最终趋向 $x=0$——完全背叛。
平均场近似是"充分混合"的世界:每个人和所有人等概率相遇。但我的格子不是。空间结构可能撑住合作。
模拟与结果
以下实验在 $50 \times 50$ 的网格上跑,每步同步更新,博弈轮数足以让系统稳定。
实验一:固定密度,看演化轨迹
设置 population density $\rho = 0.3$,变化初始合作者比例 $x_0$。
直观猜测:$x_0$ 越高,合作者撑得越久;低 $x_0$ 直接崩溃。但网格结构可能让局部高合作区在早期顶住一阵。
实际跑下来的感觉是,合作者的"存活窗口"很窄。在空间结构让局部聚集维持了一个短暂的防御期之后,D 策略终究会从边界渗透进来——一个合作团簇的外围总是最先被侵蚀。
实验二:固定初始比例,看密度效应
设置 $x_0 = 0.5$,变化 population density $\rho \in [0.02, 1.0]$,跑完后计算 $\pi_C$ 和 $\pi_D$ 的均值之差。
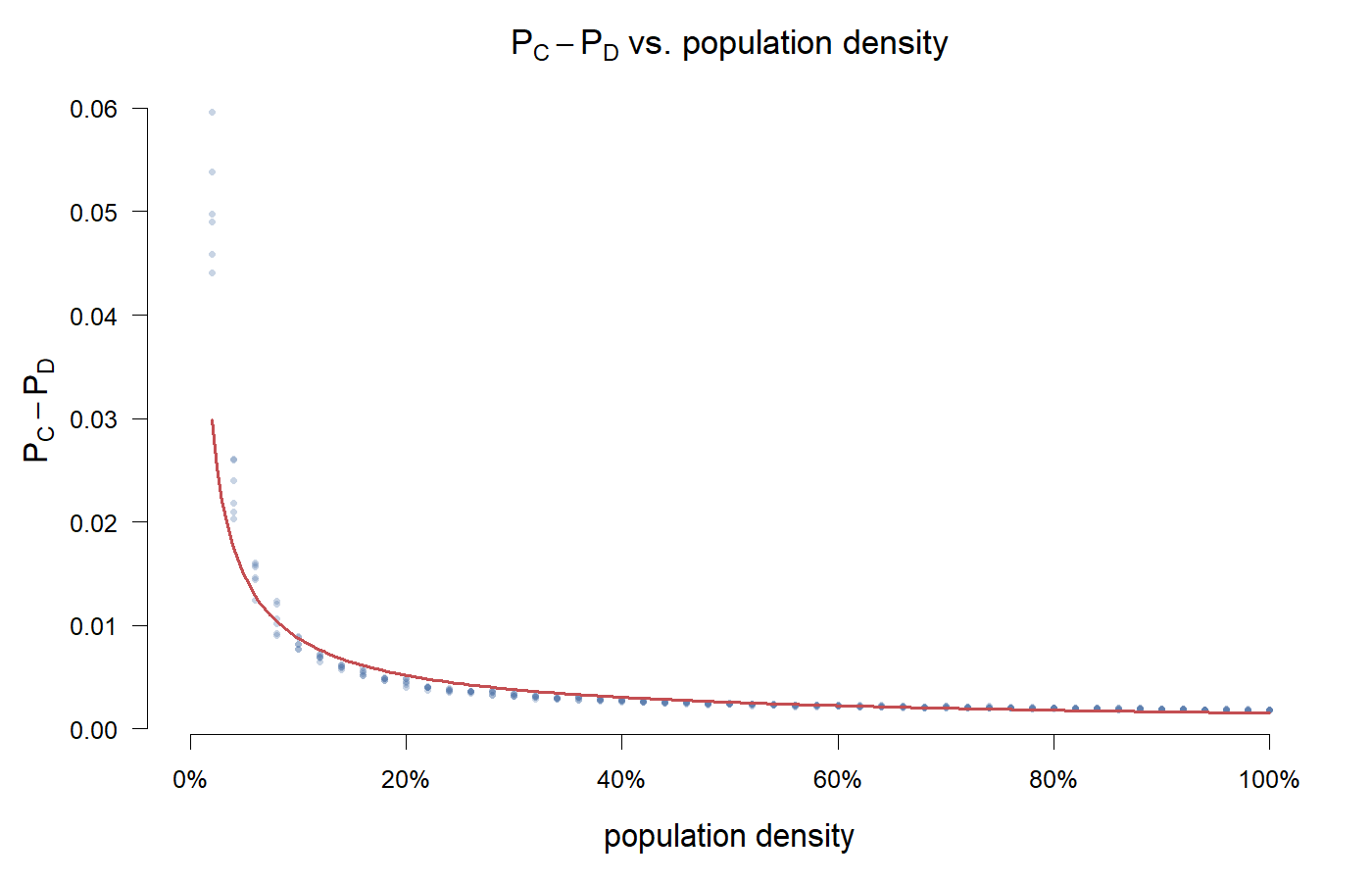
纵轴是 $P_C - P_D$,合作者的收益减去作弊者的收益。全部为正——也就是说,在所有密度下,合作者的平均收益都高于作弊者。这是在纯随机交互的平均场方程里不可能出现的东西:空间结构给 C 策略画出了一个防御地带。
但趋势一目了然。低密度时 $P_C - P_D$ 接近 0.05,高密度时掉到大约 0.002。差了 25 倍。
密度 2% 时,格子稀稀拉拉,agent 之间大部分时间是孤立的。孤立 agent 没邻居可配对——这步就没博弈、没得分波动,守住现有策略等于不亏。合作者还有偶尔撞见同类并互惠的机会。作弊者孤立的次数同样多,但孤立状态下无事发生,作弊无利可图。
密度越高,相遇越频繁。每次相遇都是一次 C 被 D 剥削的机会。即使 C-C 互惠更多,也是双方各付成本养公共池塘——刚好便宜了旁边那个什么都不贡献的 D。
Greig 的酵母实验给出了相同的方向:密度越高,作弊越有利。只不过他们是从作弊者的视角——高密度下作弊 fitness > 1。我这张图从合作者的视角——优势还在,但不断萎缩。
有意思的是,优势归零的临界密度在这组参数下始终没有出现。$P_C - P_D$ 趋近于零但没触零。可能在更极端的 payoff 设定下(比如把 T 拉更高),合作者的优势会被彻底抹掉。
(或者把 Von Neumann 换成 Moore——八个邻居比四个更容易暴露在 D 面前。还没来得及试。)
写在之后
Greig 的论文让我意识到一件事:囚徒困境的"困境"不在于无解,而在于它的解依赖于结构。相同的博弈规则,在稀疏和密集的社交网络中给出相反方向的结果。
复制子动态提供了基准预测——充分混合、只看当期收益时的演化方向。但真实系统的空间结构——无论是琼脂板上的邻居半径,还是格子上的 Von Neumann 邻域——一放进去,预测就变了。
ABM 的价值就在这里:把结构放进去,看涌现出什么。
实验一和实验二跑下来的感觉是,空间结构像一层缓冲垫——不是扭转方向,而是拖延速度。合作者注定要输给 D,但在格子上输得比平均场方程预测的要慢。
但写完之后,我越想越觉得差了一层。
Greig 的实验是在 sucrose 板上跑的。蔗糖是唯一碳源。稀疏的板子上,糖多细胞少——每个细胞周围的蔗糖约等于无限。密集的板子上,细胞挤在一起,糖很快耗光——你周围的糖取决于邻居分泌了多少酶。
我的 ABM 预设 payoff 矩阵是铁板一块。$T=5, R=3, P=1, S=0$——不管密度多少,一次博弈的收益永远不变。但酵母的 invertase 不是一台独立于环境运行的博弈机。它消化蔗糖。糖少了,消化量就少了。消化量不只取决于你分不分泌酶,还取决于蔗糖还剩多少。
如果蔗糖见底了——一群细胞挤在一起,谁也不分泌酶——双双背叛的 payoff 可能不是 $P=1$,是负数。什么都消化不了。细胞饿着。反而是合作-合作虽然要付酶的成本,起码还能分到一点糖。
也就是说,Greig 的酵母可能并不总是在玩囚徒困境。
在糖充裕的高密度区,作弊者靠剥削合作者赚饱——标准 PD。但在糖匮乏的边缘——稀疏群落耗完本地蔗糖之后——博弈的底层从囚徒困境滑向了别的东西。双双背叛不再是低收益,是负收益。合作的相对劣势突然就不见了。
策略消耗资源,资源改写矩阵,矩阵重塑策略。闭环一旦形成,囚徒困境的"困境"就不是一个给定了边界条件的优化问题——它的边界条件本身就是变量。
这还只是推测。要验证它,得把酶动力学写进 ABM——糖的浓度作为局部变量,payoff 从糖浓度里实时算出来。不是"密度决定支付矩阵",是"密度通过资源消耗间接改变支付矩阵"。
一个方向。